-
Trivial Transfer Graphene for Cryo-EM Grids - Max Planck Institute of Biophysics, 2022
May 28, 2026 | ACS MATERIAL LLCKlusch, N. et al. (2022). Cryo-EM structure of the respiratory I + III2 supercomplex from Arabidopsis thaliana at 2 Å resolution. *Nature Plants*. https://doi.org/10.1038/s41477-022-01308-6
Nature Plants · 2022
Researchers used ACS Material Trivial Transfer single-layer graphene to prepare cryo-EM grids and resolve the Arabidopsis I+III2 respiratory supercomplex at 2 Å.
About this research
Researchers at the Max-Planck-Institute of Biophysics used ACS Material's Trivial Transfer single-layer graphene as a cryo-EM grid support to determine the 2 Å resolution structure of the Arabidopsis thaliana respiratory I+III2 supercomplex, reported in Nature Plants (2022). The work, conducted with collaborators at Leibniz Universität Hannover, resolves 68 protein subunits, 94 bound cofactors, lipids and metal ions, and 4,837 ordered water molecules. The map reveals a previously undefined plant-specific complex I subunit (P9), a coenzyme A bound in the γ-carbonic anhydrase module that defines a second catalytic centre, and atomic-level water structures at the proton-exit and ubiquinone-binding sites of complex III2.
Why this research matters: mitochondrial respiratory supercomplexes channel electrons from NADH to ubiquinone and onward through cytochrome bc1, and their architecture controls cellular energy supply. Plants are unique in that their complex I carries a γ-carbonic anhydrase module potentially involved in CO2 recycling for photosynthesis, and their complex III incorporates the mitochondrial processing peptidase. Earlier negative-stain and lower-resolution cryo-EM studies of plant supercomplexes (18–20 Å, then 2.9–3.9 Å for the isolated complexes) could not resolve the small accessory subunits, lipids or water networks that stabilize the assembly. A true atomic-resolution view was needed to settle long-running questions about proton translocation pathways, Rieske head dynamics, and the role of plant-specific subunits at the I/III2 interface.
How the ACS Material product was used: Quantifoil R 1.2/1.3 400 Cu grids were cleaned in chloroform and then coated either with a 2 nm evaporated carbon layer (Leica EM ACE600) or with a graphene single layer supplied by ACS Material LLC under the Trivial Transfer label. The graphene-supported grids were pre-treated to mitigate air–water interface denaturation, functionalized with 20 µM 1-polybutyric acid, or simply glow-discharged for 15 s at 15 mA. The purified I+III2 supercomplex, isolated by digitonin solubilization and exchanged into glyco-diosgenin (GDN), was applied at 0.18 mg/mL and plunge-frozen in liquid ethane after a 4.5 s blot in a Mark IV Vitrobot operated at 10 °C and 70% humidity. The choice of graphene support was important: single-layer graphene presents an atomically thin, low-background substrate that preserves particle distribution and orientation for fragile membrane-embedded assemblies, enabling collection at 300 kV on a Titan Krios G4i.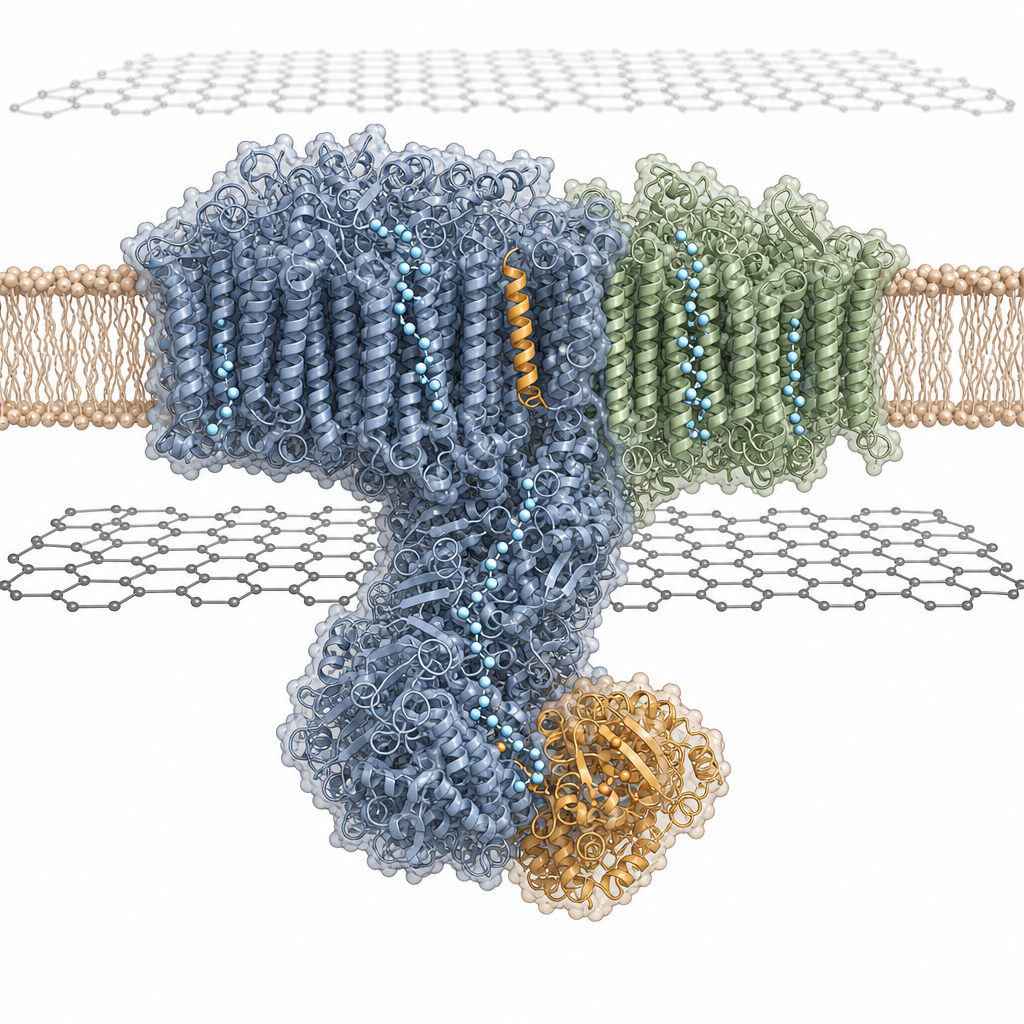
Key results: single-particle analysis yielded a 2.36 Å consensus reconstruction; particle subtraction and multibody refinement of the components improved this to 2.03 Å (1.9 Å after density modification). Two additional supercomplex conformations were resolved at up to 2.34 Å by focused 3D classification. The purified supercomplex was enzymatically active with a NADH:cytochrome c oxidoreduction activity of 2.5 U/mg. Mass spectrometry identified 48 complex I and 10 complex III subunits. The map revealed subunit B14.7 at the I/III2 interface (absent in earlier free complex I maps), reassigned MNLL15 to NUXM, and resolved the 62-residue plant-specific P9 subunit encoded by At1g67785, which spans the inner mitochondrial membrane near the ubiquinone binding site. The interface includes three contact zones; site 3 is mediated by P9 and QCR6 and is unique to plants. The angle between complex III2 and the complex I membrane arm is 65–67° (versus ~55–58° in ovine), and the membrane bends by ~6–10° around the supercomplex. The γCA2–γCAL2 interface harbours a butyryl- or crotonyl-CoA cofactor, and three potential proton-entrance half-channels (at ND2, ND4, ND5) and two lumenal exit channels were mapped. In all four Qo/Qi sites the ubiquinone is resolved, with the Rieske FeS cluster locked 6.8 Å from Qo and H237 at 2.9 Å hydrogen-bond distance.
Applications and outlook: the atomic detail extracted here informs models of proton-coupled electron transfer in mitochondrial bioenergetics, plant-specific carbon recycling via the γCA module, and supercomplex assembly more broadly. It also illustrates how graphene supports can push cryo-EM of large, lipid-rich membrane assemblies into the sub-2 Å regime, a methodology directly transferable to studies of photosystems, ATP synthases, transporters and viral envelope complexes. Follow-up work points to functional dissection of P9 and B14.7 in supercomplex stability, in vivo studies of crotonyl-CoA in plant mitochondrial metabolism, and structural comparisons with the Vigna radiata I+III2 supercomplex published in parallel.
Why this matters for researchers: structural biologists working at the resolution frontier of cryo-EM increasingly rely on monolayer graphene supports for thin-ice, low-background imaging. The Trivial Transfer single-layer graphene product used in this study is available from ACS Material to researchers preparing grids for membrane protein, supercomplex and small-particle reconstructions, where particle preservation and orientation distribution are critical for reaching atomic resolution.How ACS Material products were used
- Trivial Transfer® Graphene (single layer) (Trivial Transfer Series) — “They were coated with a 2 nm carbon layer using a Leica EM ACE600 high vacuum sputter coater or a graphene trivial transfer single layer (ACS Material LLC).”
Product Performance in this StudyThe Trivial Transfer single-layer graphene served as an alternative support film on Quantifoil cryo-EM grids to the conventional 2 nm sputter-coated carbon layer. Graphene-coated grids were functionalized with 1-polybutyric acid or glow-discharged, and supported the successful acquisition of a 2 Å resolution cryo-EM reconstruction of the Arabidopsis I+III2 supercomplex, evidencing high suitability for high-resolution single-particle cryo-EM of fragile membrane protein assemblies.
Related product categories
Frequently asked questionsWhy use single-layer graphene supports for cryo-EM grids?
Single-layer graphene is atomically thin, electrically conductive, and mechanically robust. As a cryo-EM support film it produces almost no scattering background, helps distribute particles uniformly, mitigates the air–water interface denaturation that destroys fragile complexes, and improves preferred-orientation problems. In this Arabidopsis supercomplex study, graphene-coated Quantifoil R 1.2/1.3 grids supported reconstructions at 2.03 Å, ultimately reaching 1.9 Å after density modification.
How was the graphene support prepared before sample application?
Quantifoil R 1.2/1.3 400 Cu grids were first cleaned in chloroform for one hour. A Trivial Transfer single-layer graphene film from ACS Material LLC was then transferred onto the grid. The graphene-coated grids were pre-treated against air–water interface denaturation, functionalized with 20 µM 1-polybutyric acid, or simply glow-discharged for 15 s at 15 mA before applying the 0.18 mg/mL I+III2 supercomplex sample.
What new structural insights came from the 2 Å Arabidopsis I+III2 supercomplex?
The structure resolved 68 subunits, 94 cofactors and 4,837 ordered waters. It identified a new 62-residue plant-specific complex I subunit (P9, encoded by At1g67785), located B14.7 at the I/III2 interface, revealed a butyryl- or crotonyl-CoA bound at the γCA2–γCAL2 catalytic interface, mapped three matrix proton-entrance half-channels (ND2, ND4, ND5) and two lumenal exit channels, and showed ubiquinone bound in all four Qo/Qi sites with the Rieske head locked in the b-state.